Premium USA-Made Research Compounds
Browse lab-tested peptides, research liquids, capsules and more.
Antimicrobial Peptides (AMPs): Mechanisms of Action, Research Models & Biofilm Disruption Studies
The rise of antibiotic-resistant bacteria has forced a fundamental rethinking of how researchers approach the microbiology of infection. Traditional small-molecule antibiotics — penicillins, cephalosporins, fluoroquinolones — operate by targeting specific bacterial proteins or enzymes. That specificity is, paradoxically, both their strength and their Achilles heel. Bacteria evolve resistance through mutation or gene acquisition, and specific targets become specific vulnerabilities that natural selection efficiently circumvents. The result is a well-documented global crisis of antimicrobial resistance (AMR).
Against that backdrop, antimicrobial peptides (AMPs) have attracted intensifying research interest. Not because they are new — these molecules have been part of innate immune defense for hundreds of millions of years across virtually every branch of life — but because their mechanism of action is fundamentally different. Understanding that difference, and what it means for research in 2026, is the subject of this overview.
What Are Antimicrobial Peptides? A Taxonomic Overview
AMPs are short peptides, typically 10–50 amino acids, that exhibit direct antimicrobial activity against bacteria, fungi, viruses, or parasites. They are found across the full breadth of living organisms: in frog skin (magainins, first isolated from Xenopus laevis), insect hemolymph (cecropins), mammalian neutrophils and epithelial cells (defensins, cathelicidins), and even plants and bacteria themselves. This universal distribution is itself scientifically telling — AMPs appear to represent a conserved evolutionary strategy for host defense.
Looking for Premium Research Compounds?
The major AMP classes in current research include:
- Defensins — cysteine-rich, β-sheet structured; divided into α-defensins (predominantly in neutrophil granules), β-defensins (epithelial surfaces), and θ-defensins (cyclic, found in Old World primates)
- Cathelicidins — characterized by a conserved cathelin prodomain; the most studied human representative is LL-37
- Magainins — α-helical, membrane-active peptides from amphibians; model compounds for membrane disruption research
- Cecropins — originally identified in the silkmoth; broad-spectrum activity via membrane permeabilization
- Polymyxins — cyclic, lipopeptide structure; used clinically against Gram-negative bacteria (notable as an exception where an AMP has entered clinical use)
What unites these structurally diverse molecules is a shared functional logic: selective cytotoxicity toward prokaryotic membranes, mediated predominantly by electrostatic interactions.
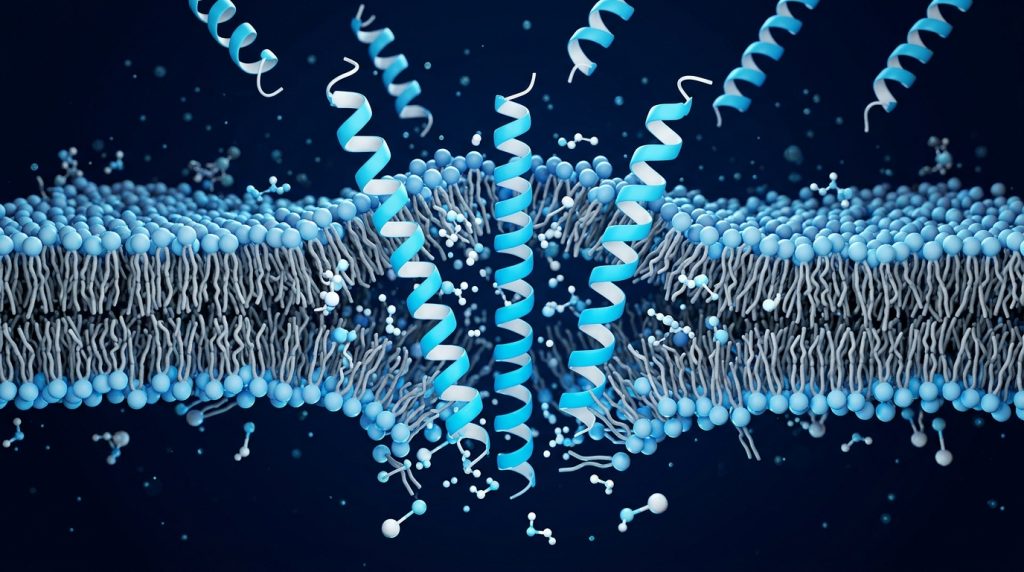
Membrane Disruption: The Mechanistic Core
Why are bacteria selectively targeted? The answer starts with membrane composition. Bacterial membranes carry net negative surface charges, arising from lipopolysaccharides in Gram-negative outer membranes and phosphatidylglycerol or cardiolipin in Gram-positive surfaces. Most AMPs are cationic — they carry a net positive charge at physiological pH. Electrostatic attraction between cationic AMP and anionic bacterial membrane drives the initial association.
Eukaryotic cell membranes, by contrast, present a largely neutral outer leaflet dominated by phosphatidylcholine and sphingomyelin. Cholesterol further rigidifies and stabilizes mammalian membranes in ways that reduce AMP insertion efficiency. This charge and structural differential is the molecular basis of selectivity — and it is why AMPs preferentially disrupt bacterial membranes rather than host cell membranes, at least within a defined concentration range.
Once associated with the bacterial surface, AMPs can disrupt membrane integrity through several structural mechanisms, which researchers have classified using biophysical models:
- Carpet model — AMPs accumulate on the membrane surface in an oriented monolayer until a critical threshold is reached, at which point the membrane dissolves in a detergent-like fashion
- Toroidal pore model — AMPs insert into the bilayer and induce a continuous curvature, forming pores lined by both peptide and lipid headgroups (magainins exemplify this mechanism)
- Barrel-stave model — AMPs insert perpendicularly into the bilayer and oligomerize to form transmembrane channels, with peptides forming the barrel staves around a central aqueous pore
In practice, many AMPs do not conform exclusively to one model; the operative mechanism may depend on membrane composition, peptide concentration, and lipid-to-peptide ratios. Distinguishing mechanisms experimentally — using techniques such as oriented circular dichroism, solid-state NMR, or cryo-EM of model membranes — remains an active research area.
LL-37: The Most-Studied Human Cathelicidin
Of all AMPs in the research literature, LL-37 occupies a unique position. It is the only cathelicidin found in humans, cleaved from the hCAP18 precursor protein and expressed primarily in neutrophils, monocytes, natural killer cells, and epithelial cells of the skin, lung, and gut. Its structural and functional characterization has proceeded in extraordinary detail.
LL-37 is a 37-amino-acid α-helical peptide with broad-spectrum antimicrobial activity. But what elevates it beyond a simple membrane-disruptor is its immunomodulatory profile. LL-37 binds to FPRL1 (formyl peptide receptor-like 1, also known as FPR2) on immune cells, triggering chemotaxis, calcium mobilization, and modulation of inflammatory signaling. It also binds extracellular DNA and LPS, activities with implications for biofilm research and inflammatory response modulation respectively.
Researchers studying LL-37 have characterized its role in KAMP (Kidney AMP) biology — where cathelicidin expression in the renal tubular epithelium appears to serve localized antimicrobial and regulatory functions. This tissue-specific expression work has expanded understanding of how AMPs function not just as systemic defense molecules but as locally regulated effectors at distinct epithelial interfaces.
Biofilm Disruption: A Research Frontier
Biofilms represent one of the most clinically and scientifically intractable problems in microbiology. When bacteria transition from planktonic (free-floating) to biofilm states, they embed themselves in a self-produced extracellular matrix — a complex mixture of polysaccharides, proteins, eDNA (extracellular DNA), and lipids. This matrix is not merely structural; it creates diffusion barriers to antimicrobials, provides pH buffering against metabolic byproducts, and facilitates phenotypic heterogeneity within the bacterial population. Biofilm-associated bacteria can be 10–1,000 times less susceptible to conventional antibiotics than their planktonic counterparts.
AMPs present a mechanistically distinct approach to this problem. Several mechanisms contribute to AMP activity against biofilms:
First, the cationic nature of many AMPs promotes electrostatic binding to the anionic components of the extracellular matrix itself — particularly eDNA and certain polysaccharides — disrupting matrix architecture. Second, small AMP size (relative to most antibiotics) may improve diffusion through matrix barriers. Third, membrane-active AMPs retain activity against metabolically dormant cells in deeper biofilm layers, where many conventional antibiotics that require active metabolic processes lose efficacy.
Research models for biofilm disruption include static biofilm formation assays (crystal violet staining for quantification), drip-flow reactor models for dynamic biofilm growth, and confocal laser scanning microscopy with fluorescent labeling to visualize three-dimensional biofilm architecture before and after AMP exposure. Minimum biofilm eradication concentration (MBEC) assays, analogous to MIC assays for planktonic cells, provide standardized comparative data.
Synthetic AMP Design: 2026 Research Trends
Natural AMPs face well-characterized limitations in research and potential applied contexts: susceptibility to protease degradation in biological matrices, significant serum protein binding that reduces effective concentration, and cytotoxicity to eukaryotic cells at concentrations above the therapeutic window. These challenges have driven extensive interest in synthetic AMP design — engineering molecules that retain or improve antimicrobial activity while addressing these liabilities.
Several strategies have emerged as the leading approaches in 2026:
D-amino acid substitution replaces one or more L-amino acids with their mirror-image D-enantiomers. Since proteases are stereospecific and operate almost exclusively on L-amino acid substrates, D-substituted AMPs demonstrate dramatically improved proteolytic stability while retaining membrane-disruptive activity (membranes being achiral at the level of gross biophysical interaction).
Cyclization — connecting the N- and C-termini, or forming side-chain crosslinks — increases conformational rigidity, protease resistance, and in some cases membrane affinity. θ-defensins, the only naturally cyclic mammalian AMPs (silenced in humans by a premature stop codon), have served as structural templates for this design strategy.
Lipid conjugation appends fatty acid chains to AMP scaffolds, enhancing membrane affinity and antimicrobial potency — though careful optimization is required to avoid increased cytotoxicity against eukaryotic membranes, which can also be perturbed by highly lipophilic compounds.
Computational approaches — molecular dynamics simulation of AMP-membrane interactions, machine learning models trained on large AMP activity datasets — are increasingly guiding rational design, reducing the empirical screening burden that characterized earlier generations of synthetic AMP work.
Research Limitations and Open Questions
Scientific rigor requires acknowledging what AMPs cannot yet do in research models. Serum protein binding remains a significant confounder in translating in vitro minimum inhibitory concentration (MIC) data to in vivo models — proteins in biological fluids sequester AMPs and reduce their effective free concentration substantially. Cytotoxicity windows are narrower than for many conventional agents, requiring precise concentration control in cell-based assays. And while resistance development to AMPs is generally considered harder to evolve (for mechanistic reasons discussed above), it is not impossible — bacterial resistance mechanisms including proteolytic degradation, membrane remodeling, and electrostatic charge modification have been documented under selection pressure.
These are research challenges, not research-ending obstacles. They define the experimental parameters that rigorous AMP researchers must characterize and account for.
Conclusion
Antimicrobial peptides occupy a scientifically distinctive position in the landscape of anti-infective research. Their evolutionary antiquity, mechanistic diversity, and physical mode of action against bacterial membranes make them irreplaceable research tools for understanding innate immunity, biofilm biology, and the cellular biophysics of membrane disruption. As synthetic design approaches mature and as resistance mechanisms are mapped in greater detail, AMP research is increasingly well-positioned to contribute foundational knowledge to one of the most urgent problems in modern microbiology.
For Research Purposes Only: The information presented in this article is intended solely for scientific research and educational purposes. These compounds are not approved for human use and should only be handled by qualified researchers in appropriate laboratory settings in compliance with all applicable regulations.
Continue Your Research
Explore our complete catalog of premium research compounds.