Premium USA-Made Research Compounds
Browse lab-tested peptides, research liquids, capsules and more.
For research use only. Not for human consumption or veterinary use.
Here’s a situation every somatotropic researcher eventually runs into: three compounds on the shelf, all of them secretagogues, all hitting GHSR-1a — and the protocol decision matters more than most people initially realize.
Ipamorelin came in 1998. GHRP-6 goes all the way back to 1980. GHRP-2 landed in the early 80s as researchers tried to refine what Bowers had built at Tulane. Different eras, different structural logic, different receptor fingerprints. Calling them interchangeable because they share a primary target is like saying a scalpel and surgical scissors are the same tool because they both cut. The overlap is real. So are the differences — and in practice, picking the wrong one doesn’t just add noise; it can invalidate the question the protocol was designed to answer.
Looking for Premium Research Compounds?
This piece works through all of it: mechanism, selectivity, off-target signaling, pulsatile GH dynamics, and where each compound actually earns its place in a well-designed protocol.
What Are GHRPs? The Growth Hormone Secretagogue Mechanism
Growth hormone-releasing peptides are synthetic compounds that mimic ghrelin — a 28-amino-acid gut peptide that drives GH secretion by binding the growth hormone secretagogue receptor type 1a (GHSR-1a). That receptor lives in two strategically important locations: the hypothalamus and the anterior pituitary. Both matter for understanding how GHRPs actually work.
The mechanism isn’t as simple as “compound binds receptor, GH comes out.” GHRPs amplify an existing pulsatile signal rather than generating one from nothing. The hypothalamus is already producing rhythmic GHRH pulses; what GHSR-1a agonism does is sensitize pituitary somatotrophs to those incoming signals, making each pulse more forceful. The downstream sequence — Gαq/11 coupling, phospholipase C activation, IP3-mediated calcium mobilization, GH exocytosis — is the same across all three compounds. What differs is where else they reach.
Stack any GHRP with exogenous GHRH and the GH pulse climbs sharply — several-fold above what either agent produces alone. That’s a genuine synergy, not additive noise, and it’s why GHRH/GHRP combination protocols have become standard in secretagogue research. The three compounds all preserve that synergy. They diverge on nearly everything else.
Pulsatile GH Release: Why It Matters
GH fires in pulses — roughly 6 to 12 per day in humans, with the largest occurring during slow-wave sleep. That architecture isn’t incidental. Pulse amplitude and frequency drive downstream IGF-1 synthesis, drive lipolysis, determine anabolic signaling in muscle and bone. Continuous, non-pulsatile GH elevation is actually a problem: sustained receptor activation leads to desensitization and blunted response. GHRPs amplify the existing rhythm rather than steamrolling it — which is one reason they’ve become preferred tools over exogenous GH in mechanistic studies where physiological signal fidelity matters.
In rodent models, GH pulses occur every 3–4 hours with near-zero interpulse troughs. Between pulses, somatostatin tone is high. Administer a GHRP during a high-somatostatin window and the GH response is blunted — exactly what the physiology predicts from a compound that respects normal secretory architecture. That behavioral validation matters when interpreting study results.
Ipamorelin: The Selective Secretagogue
Novo Nordisk developed Ipamorelin in the mid-1990s; the first full characterization appeared in 1998. Structurally it’s a pentapeptide — Aib-His-D-2Nal-D-Phe-Lys-NH₂ — and those non-standard residues at positions 1 and 3 (alpha-aminoisobutyric acid and D-beta-naphthylalanine) turn out to be doing most of the pharmacological work. They’re what give Ipamorelin its clean selectivity profile. Without them, the off-target activity looks a lot more like GHRP-2.
Mechanism of Action
Ipamorelin is a selective GHSR-1a agonist. That word — selective — isn’t marketing language here; it’s the mechanistic observation that defines everything about how this compound behaves in a study. It hits the receptor, runs the Gαq/11 → PLC → IP3 → calcium cascade, delivers a robust GH pulse, and essentially stops there. Corticotrophs don’t get appreciably activated. Lactotrophs stay quiet. The Ankersen et al. characterization work established this pattern clearly at near-maximal GH-stimulating concentrations — no statistically significant plasma ACTH, cortisol, or prolactin elevation. GHRP-2 at comparable concentrations elevated cortisol. GHRP-6 showed intermediate cortisol activity. Ipamorelin didn’t.
Why that selectivity exists at the receptor level isn’t entirely settled — it may reflect biased agonism, where the specific binding geometry favors the somatotroph signaling pathway over the corticotroph one, even though both cell populations express GHSR-1a. The practical consequence is clear regardless of the mechanistic explanation: robust GH secretion without the hormonal background noise that complicates interpretation.
Selectivity Profile
The cortisol advantage is the headline, but there are a few others worth noting. Ipamorelin shows less receptor desensitization than some comparable compounds in extended preclinical protocols — GH responsiveness holds up over time in a way that matters for studies running weeks rather than days. Prolactin stays minimal throughout. And the appetite effects that define GHRP-6’s profile? Essentially absent at GH-relevant concentrations.
For studies where cortisol or prolactin elevation would confound the primary readout — most mechanistic GH studies — that cleanliness isn’t a minor methodological preference. It’s the entire reason to choose Ipamorelin over the alternatives.
Research Applications
The clean signal profile makes Ipamorelin the default choice in several active research areas:
- GH deficiency models: Restoring pulsatile GH in hypophysectomized or GH-deficient rodents without cortisol confounders altering metabolic readouts
- Bone biology: Preclinical work examining GH’s direct effects on osteoblast activity and trabecular bone formation — where baseline cortisol elevation would directly interfere with the question
- Muscle biology: GH-mediated satellite cell activation, myofibrillar protein synthesis, and recovery dynamics in animal models
- Aging/somatopause research: Restoring GH pulsatility in aged rodent cohorts where baseline HPA reactivity is already elevated — adding cortisol signal on top would make interpretation untenable
- Sleep architecture studies: Correlating GH pulse timing with slow-wave sleep stages; prolactin is an active confounder in sleep biology, making Ipamorelin’s minimal prolactin response a genuine asset
Novo Nordisk advanced Ipamorelin into Phase II investigational work for postoperative ileus before halting the program — for business reasons, not because the compound failed pharmacologically. The early human pharmacology data remains in the published record and continues to inform preclinical-to-translational research frameworks.
GHRP-2: The Potent Secretagogue
GHRP-2 is D-Ala-D-β-Nal-Ala-Trp-D-Phe-Lys-NH₂. Hexapeptide. Came out of the early GHRP-6 optimization work in the 1980s as researchers systematically modified the lead structure looking for improved potency and stability. They found it. By raw GH-release metrics, GHRP-2 is the most potent of these three compounds. The tradeoff, and it’s a real one, is HPA axis co-activation.
Mechanism of Action
GHRP-2 is a full GHSR-1a agonist with higher binding affinity than GHRP-6 in most competitive displacement assay formats. The primary signaling cascade is identical to Ipamorelin’s — Gαq/11 coupling, PLC, IP3, calcium, GH exocytosis. But GHRP-2 adds something Ipamorelin doesn’t: direct suppression of hypothalamic somatostatin release. Somatostatin is the brake pedal on GH secretion. GHRP-2 is simultaneously pressing the accelerator (GHSR-1a activation) and releasing the brake (somatostatin inhibition). That’s why peak GH values under GHRP-2 run higher than under Ipamorelin at equivalent experimental doses — roughly 30–50% higher pulse amplitude in most head-to-head rodent studies, though the specific number varies with model and dose.
The HPA co-activation mechanism runs through CRH. GHSR-1a activation in the hypothalamus triggers CRH release, which then hits anterior pituitary corticotrophs, producing ACTH and downstream cortisol elevation. The cortisol response tracks roughly proportionally with GH pulse amplitude — bigger GH pulse, bigger cortisol co-secretion. Bowers’ early human pharmacology work in the 1990s documented this clearly.
Selectivity Profile
GHRP-2 produces the highest cortisol and prolactin co-secretion of the three compounds. That’s not a design flaw in the compound — it’s a characteristic that shapes when it’s the right tool. Metabolic research in obese animal models, for instance, sometimes benefits from the corticotrophic co-activation as an additional readout. Cardiac research using GHRP-2’s cardioprotective effects doesn’t typically require cortisol isolation. The problem arises when researchers default to GHRP-2 because of its high potency, then struggle to separate GH effects from the cortisol signal riding alongside.
One finding worth highlighting: repeated GHRP-2 exposure maintains GH responsiveness better than several synthetic GHRH analogues in long-duration preclinical protocols. Receptor desensitization is less of a concern here than intuition might suggest.
Research Applications
- Maximum GH pulse amplitude studies: When peak somatotroph stimulation is the primary endpoint, GHRP-2 delivers values the other two can’t match
- IGF-1 axis research: Robust and sustained GH elevation drives sustained IGF-1 upregulation — useful in models tracing the complete GH→IGF-1→tissue signaling cascade
- Metabolic models: Body composition, adipose tissue lipolysis, and insulin sensitivity studies in diet-induced obesity where HPA co-activation is either controlled for or informative
- GHSR-1a pharmacology: Receptor characterization in knockout or mutant models where potent full-agonist engagement is needed to saturate the receptor
- Cardiac research: Preclinical evidence for GHSR-1a-mediated cardioprotective effects — potentially through PI3K/Akt survival signaling in cardiomyocytes, independent of GH elevation
GHRP-6: The Original Ghrelin Mimetic
GHRP-6 is the compound that started all of this. His-D-Trp-Ala-Trp-D-Phe-Lys-NH₂. Cyril Bowers at Tulane synthesized it in 1980 as a derivative of met-enkephalin opioid peptides — not, interestingly, because he was trying to build a GH secretagogue. The GH-stimulating activity was essentially a discovery. What followed was nearly two decades of research chasing the receptor that GHRP-6 was activating, a pursuit that eventually led to the identification of ghrelin as the endogenous GHSR-1a ligand in 1999. The causal chain runs directly: no GHRP-6 in 1980, no ghrelin in 1999. That’s the kind of historical weight this compound carries.
Mechanism of Action
GHRP-6 is a full GHSR-1a agonist and, structurally, the closest analogue among these three to endogenous ghrelin. Ghrelin has a unique n-octanoyl modification at Ser3 that’s responsible for much of its receptor potency; GHRP-6 achieves comparable activation through D-Trp and Trp residues. The mechanistic consequence is that GHRP-6 activates the full biological program that ghrelin controls — not just GH secretion, but somatostatin suppression, appetite signaling via NPY/AgRP pathways, gastric motility, and elements of the reward circuitry. That’s a broad experimental footprint. For some studies it’s exactly what’s needed. For others, it creates an interpretation problem.
Appetite Stimulation: The Ghrelin Effect
This is GHRP-6’s most distinctive characteristic — and the one most likely to catch researchers off guard the first time they run it. Ghrelin is the primary orexigenic hormone. It drives feeding behavior between meals, promotes NPY and AgRP release in the arcuate nucleus, and upregulates adipose tissue storage. GHRP-6 activates those same circuits reliably and at doses well below those needed for maximal GH stimulation.
Rodent studies consistently document 40–60% increases in food intake following GHRP-6 administration, depending on model and dose. GHRP-2 shows appetite stimulation too, but weaker and less consistent. Ipamorelin produces essentially none at GH-relevant concentrations. So if the research question involves the somatotropic axis and only the somatotropic axis — pure GH biology — GHRP-6’s orexigenic activity is noise. But if the question extends to energy homeostasis, feeding behavior, or the interplay between GH and metabolic state, that appetite signal becomes the most valuable variable in the study.
Selectivity Profile
Cortisol and prolactin co-secretion with GHRP-6 sits between Ipamorelin (minimal) and GHRP-2 (highest). The HPA activation is present but typically less pronounced in head-to-head studies. What GHRP-6 contributes that neither of the other compounds can replicate is the gastric motility component — GHRP-6 accelerates gastric emptying through ghrelin receptor activation in gut vagal afferents, a pathway that’s both a potential confounder in some study designs and the entire point in others. Gastric dysmotility research, gut-brain axis studies, and energy balance models where ghrelin’s full biological program needs to be active — those are the use cases where GHRP-6’s breadth is an asset rather than a liability.
Research Applications
- Appetite and feeding behavior: The established primary application — characterizing NPY/AgRP-mediated orexigenic signaling in preclinical models using the compound most reliably activating those circuits
- Energy homeostasis: Ghrelin’s role in energy balance, adipogenesis, and metabolic adaptation during caloric restriction; GHRP-6 activates the full program
- Gastric motility: Accelerated gastric emptying via vagal ghrelin receptors — directly applicable in dysmotility research
- Historical/reference work: Decades of foundational GH secretagogue literature used GHRP-6 as the reference compound; researchers building on or replicating that work need the same tool
- Cardiac and cytoprotective studies: Ischemia-reperfusion injury models have accumulated preclinical evidence for GHRP-6-mediated cardioprotection — potentially through GHSR-1a pathways in cardiomyocytes and inflammatory modulation
- CNS/reward circuitry: GHSR-1a forms heterodimers with dopamine D1 receptors; GHRP-6’s structural proximity to ghrelin makes it the preferred tool compound for probing these non-GH CNS functions
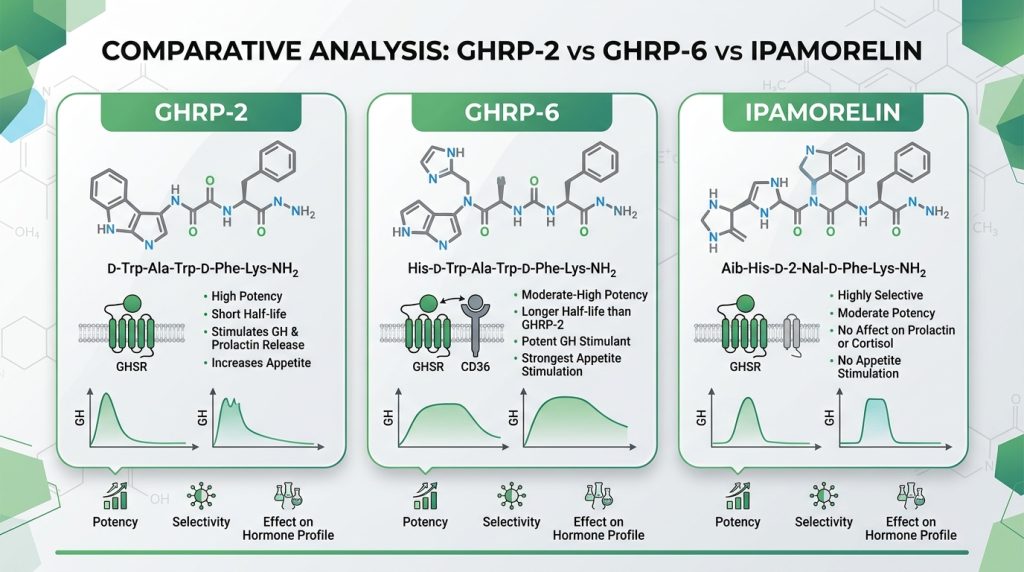
Side-by-Side Research Comparison
| Parameter | Ipamorelin | GHRP-2 | GHRP-6 |
|---|---|---|---|
| Compound class | Synthetic pentapeptide | Synthetic hexapeptide | Synthetic hexapeptide |
| Primary receptor | GHSR-1a (selective) | GHSR-1a (full agonist) | GHSR-1a (full agonist) |
| GH release potency | Moderate–High | Very High | High |
| Cortisol co-secretion | Minimal / None | Significant | Moderate |
| Prolactin co-secretion | Minimal / None | Moderate | Moderate |
| Appetite stimulation | Minimal | Moderate | Strong (NPY/AgRP mediated) |
| Somatostatin suppression | Indirect (via GHSR-1a) | Strong (direct + indirect) | Moderate |
| Gastric motility effect | Minimal | Minimal | Pronounced (ghrelin mimicry) |
| Ghrelin receptor homology | Partial mimetic | Partial mimetic | Closest structural analogue |
| Pulsatile GH preservation | Excellent | Good | Good |
| Research selectivity | Highest (GH-specific studies) | Moderate (GH + metabolic) | Broadest (GH + ghrelin axis) |
| First described | 1998 | 1984 | 1980 |
| Primary research utility | Clean GH studies, bone/muscle, aging | Max GH stimulation, IGF-1, metabolic | Ghrelin axis, appetite, gastric biology |
Selectivity Profiles in Depth
GHSR-1a Binding Affinity
All three bind GHSR-1a with high affinity — but not identically, and binding affinity doesn’t cleanly predict in vivo GH output. GHRP-2 generally shows the highest affinity in competitive displacement assays; Ipamorelin follows closely; GHRP-6 tends to come in somewhat lower depending on assay format. That said, in vivo potency ranking doesn’t track directly with Ki values. Receptor reserve, endogenous somatostatin tone at the moment of administration, and GHRH background activity all shape the actual GH pulse. This is one reason head-to-head binding assays don’t always match head-to-head secretion experiments — and why researchers occasionally get surprised when GHRP-6 outperforms GHRP-2 in certain model contexts despite lower receptor binding affinity on paper.
Cortisol and HPA Axis Interaction
The cortisol divergence between these three is the most practically significant selectivity difference — and the one most worth spending time on. GHRP-2 reliably elevates plasma ACTH and cortisol in both rodent models and the early human pharmacology studies from the 1990s. The mechanism: hypothalamic GHSR-1a activation triggers CRH release → pituitary corticotroph activation → ACTH → adrenal cortisol production. The size of that cortisol response is roughly proportional to the GH pulse amplitude.
Ipamorelin binds the same receptor and produces a comparable GH pulse — but essentially none of that cortisol cascade follows. Why? The most likely explanation is biased agonism: Ipamorelin’s specific binding conformation shifts signaling toward the somatotroph pathway rather than the corticotroph pathway, even though both cell populations express GHSR-1a. The receptor is the same; the intracellular signal balance is different. GHRP-6 lands between the two — cortisol elevation present but consistently less pronounced than GHRP-2 in head-to-head work.
The bottom line for researchers: if GH effect isolation is part of the experimental logic, and it should be in most mechanistic studies, Ipamorelin’s cortisol profile isn’t a minor convenience. It’s a methodological necessity.
Prolactin Response
Prolactin co-secretion runs the same ranking — GHRP-2 highest, GHRP-6 intermediate, Ipamorelin minimal. Arvat et al. (1999) demonstrated this pattern in controlled studies using equivalent mass doses. Prolactin is a confounder in breast tissue biology, reproductive studies, certain immune function research, and sleep architecture work. The choice of secretagogue shapes which of those variables stays quiet — and in study designs where prolactin is part of the background biology, the compound selection has direct consequences for interpretation.
The GHRH Synergy Effect
All three compounds preserve synergy with exogenous GHRH — that’s not in question. Combine any of them with CJC-1295 or another GHRH analogue and GH pulse amplitude climbs several-fold above what either agent produces in isolation. The mechanism is mechanistically additive: GHRH directly stimulates somatotrophs through a separate receptor, while GHRPs suppress somatostatin and prime GHSR-1a simultaneously. Where absolute pulse values land under combination protocols still tracks each compound’s intrinsic potency — GHRP-2 combinations produce higher peak values than Ipamorelin combinations — but the synergy architecture is preserved across all three.
How Researchers Choose Between These Compounds
The selection logic is worth making explicit rather than leaving implicit.
Choose Ipamorelin When:
- The study needs GH elevation without HPA axis activation — cortisol and prolactin need to remain as clean controls
- Long-duration protocols require sustained receptor sensitivity (Ipamorelin shows better maintenance of GH responsiveness over extended preclinical timelines)
- Investigating GH-specific effects on bone mineral density, lean tissue accretion, or sleep in clean model systems
- Working with aged animal models where baseline HPA reactivity is already elevated and adding corticotrophic stimulation would confound every readout
- Running GHRH combination protocols — Ipamorelin paired with CJC-1295 has become one of the most-studied combination protocols for clean, sustained IGF-1 elevation in animal models
Choose GHRP-2 When:
- Maximum GH pulse amplitude is the primary endpoint — nothing in this class delivers higher peak somatotroph stimulation
- The research design actively benefits from HPA co-activation as an additional readout, or controls for it directly
- Metabolic endpoints dominate — adipose lipolysis, body composition shifts, insulin sensitivity in diet-induced obesity models
- GHSR-1a pharmacology characterization in receptor-mutant or knockout models demands potent full-agonist saturation
- Studying cardiac or cytoprotective mechanisms that may run through GHSR-1a-mediated PI3K/Akt survival signaling
Choose GHRP-6 When:
- The research question extends beyond the pituitary into ghrelin axis biology — orexigenic signaling, NPY/AgRP pathways, hypothalamic feeding circuits
- Gastric motility or gut-brain axis work is part of the study design
- Replicating or building directly on the decades of foundational GHRP literature — GHRP-6 is the original reference compound and methodology comparison often demands the same tool
- Energy homeostasis models where ghrelin’s complete biological program needs to be active simultaneously
- CNS function studies probing GHSR-1a heterodimer activity with dopamine receptors — GHRP-6’s structural proximity to ghrelin makes it the right probe for that work
Pulsatile GH Release Dynamics: A Deeper Look
How GHRPs shape pulsatile GH secretion is worth understanding beyond the headline mechanism — particularly because it determines whether experimental results are physiologically interpretable or not.
Between pulses, somatostatin tone is high and GH output is near zero. Administering a GHRP during that somatostatin-dominant window produces a blunted response — which is exactly what the physiology predicts, and exactly what researchers should want to see as a validation signal. Flat GH responses to GHRP at all time points regardless of pulse phase suggest something about the HPA axis or model state is off.
Ipamorelin preserves the pulsatile architecture most faithfully. The GH signal is clean, the interpulse troughs stay low, and the response-to-administration ratio remains predictable over extended protocol durations. GHRP-2 adds somatostatin suppression, which produces larger absolute pulse amplitudes but also compresses the natural trough — the pulsatile signal becomes somewhat more continuous under repeated GHRP-2 administration. GHRP-6’s NPY/AgRP activation introduces a variable that can independently modulate GH pulse frequency in certain model contexts — a subtle but real confound in studies where pulse frequency, not just amplitude, is a primary endpoint.
Current 2025–2026 Research Landscape
The GHRP field is mature but not static. A few active directions are worth tracking.
Senescence and Aging Models
Somatopause — the progressive decline in GH pulsatility that begins in the third decade and accelerates with age — remains one of the most active areas of secretagogue research. The question researchers keep returning to is whether restoring GH pulsatility in aged subjects reverses measurable functional decline in muscle, bone, and metabolic parameters, or whether the somatopause itself reflects a broader neuroendocrine recalibration that GH restoration can’t fully address.
Ipamorelin has become the dominant tool compound in this area. The rationale follows directly from the selectivity profile: aged animal models have elevated baseline HPA reactivity, and running GHRP-2’s cortisol co-activation on top of that creates an impossible interpretation problem. Studies through 2024 continue mapping GH restoration’s effects on physical function and body composition in aged rodent cohorts using Ipamorelin as the reference secretagogue.
Cardioprotection Research
Both GHRP-2 and GHRP-6 have accumulated preclinical evidence for cardioprotective effects in ischemia-reperfusion injury models — reduced infarct size, preserved left ventricular function, attenuated inflammatory markers. Whether those effects run through GH→IGF-1 signaling, direct GHSR-1a activation in cardiomyocytes, or some combination remains an active question. GHSR-1a expression in cardiac tissue is now well-established; the downstream effectors — PI3K/Akt survival signaling, NF-κB inflammatory modulation — are being characterized. This is one of the more translatable research directions to emerge from the GHRP literature.
GHSR-1a as a Multifunctional Target
The receptor isn’t just a GH-release switch. GHSR-1a is expressed in brain regions governing reward, memory consolidation, and food-seeking behavior — areas well outside the hypothalamic-pituitary axis. A 2023 review in Frontiers in Neuroendocrinology highlighted heterodimer formation between GHSR-1a and dopamine D1 receptors as a potentially significant mechanism for ghrelin’s CNS effects. GHRP-6, as the structural ghrelin closest-analogue, has become the preferred research tool for probing these non-GH functions. The emerging picture is that GHSR-1a is a far broader target than early GH-focused research appreciated — and GHRP-6’s place in the toolkit has expanded accordingly.
Summary: Three Tools, Three Research Profiles
The underlying receptor is shared. The compounds aren’t.
Ipamorelin is precision. Clean GH signal. Minimal endocrine noise. Preserved pulsatile architecture. The best choice when the study demands GH in isolation — and when the data needs to be interpretable without cortisol or prolactin corrections running in parallel.
GHRP-2 is amplitude. Highest peak GH values in this class. Strong IGF-1 responses. HPA co-activation that’s a legitimate methodological problem in some study designs and an additional readout in others. When maximum somatotroph stimulation is the endpoint, nothing in this group matches it.
GHRP-6 is breadth. Forty-plus years of foundational literature. Appetite signaling, gastric motility, hypothalamic feeding circuits, CNS ghrelin biology — none of those research questions are accessible through Ipamorelin or GHRP-2 the way they are through GHRP-6. The compound that made ghrelin’s discovery possible remains the right tool when the question extends beyond the pituitary.
Match compound to question. It’s the first principle of secretagogue research — and these three compounds exist in the literature precisely because no single one answers all of the questions.
All compounds referenced in this article are for research use only. Information presented here is for educational and scientific purposes and does not constitute guidance for human use.
Continue Your Research
Explore our complete catalog of premium research compounds.